



pro@bio-group.net

Обзор научной литературы про слепняков травяных Лигусов, вызывающих гормональный дисбаланс и изменение вегетативного роста растений: снижение роста стеблей и увеличение роста боковых ветвей, а также деформацию плодов и семян.
Клопы-слепняки рода Lygus Hahn, 1833 относятся к трибе Травяных клопиков Mirini Hahn, 1831; к подсемейству Mirinae Hahn, 1831 [1]; к семейству Слепняков Miridae Hahn, 1831.
Семейство Слепняки (Miridae) входит в состав надсемейства Мирообразные (Miroidea), инфраотряда Цимикоморфные клопы (Cimicomorpha Leston, Pendergrast et Southwood, 1954), подотряда Клопы (Heteroptera), отряда Полужесткокрылые (Hemiptera Latreille, 1810), подкласса Насекомые (Insecta), класса Шестиногие (Hexapoda).
На Земле зарегистрировано 164 номинальных видов рода Lygus, из них в Палеарктике – 21 вид [2, 3], в Европе – 7 видов [3]:
Только первые два вида, являясь полифагами, способны проникать в тепличные сооружения, где повреждают овощные и цветочные культуры. При этом Lygus rugulipennis – не только самый распространенный, но и наиболее массовый и вредоносный вид [4].
Lygus rugulipennis Poppius, 1911 – клопик опушённый (= травяной клоп; European tarnished plant bug)
Базионим (название, данное виду при его первоначальном описании): Lygus rugulipennis Poppius, 1911 [5] (ruga=морщина, penna=крыло, т.е. сморщеннокрылый). Впервые вид описан Поппиусом по материалу из Финляндии.
Известные синонимичные названия:
Валидное название — Lygus rugulipennis Poppius, 1911.
Согласно Code EPPO (European and Mediterranean Plant Protection Organization) клопик опушённый Lygus rugulipennis кодируется как LYGURU.
Lygus rugulipennis имеет Голарктический ареал. В Неарктике (Северная Америка) распространён от Аляски до севера Калифорнии и юга Колорадо [6]. В Палеарктике вид отмечен от Великобритании и Испании до Дальнего Востока России и Японии [7, 8].
На территории европейской части России L.rugulipennis распространен на север до Мурманской и Архангельской областей и Республики Коми, на юг до Краснодарского и Ставропольского краев [2].
Взрослые L.rugulipennis средних размеров с уплощенным овальным телом (рис. 1). Длина тела самок 5,7 мм, самцов 4,7 мм. Тело покрыто тонким пушком. Окраска тела зависит от пола и сезона года. Основными цветами общей окраски являются серо-зеленый, бурый или черноватый. Обычно самки окрашены светлее самцов. Переднеспинка и надкрылья более менее явственно пунктированы.
Голова почти вертикальная, спереди выглядит более или менее треугольной; часто (особенно у самцов) с тёмно-бурым, рыжим или тёмно-красным изменчивым рисунком. Задний край темени с отчетливым поперечным рёбрышком. Лоб гладкий, без бороздок, блестящий. Верхняя половина наличника не прикрыта щеками, хорошо видна сбоку.
Глаза крупные. Внутренний край глаза сильно вогнутый.
Усики тонкие, длиннее половины тела, густо опушены короткими серебристыми волосками; прикреплены несколько выше нижнего края глаз. 3-й членик усиков короче 2-го. Длина второго членика усика больше ширины головы.
Переднеспинка трапециевидная, передний край ее явственно отделен глубокой поперечной бороздкой в виде узкого выпуклого шейного кольца. Переднеспинка в передней части с двумя небольшими слабо выступающими разъединенными мозолистыми возвышениями. Задний край переднеспинки в 1,6-2,3 раза шире головы.
Щиток треугольный. Его основание целиком прикрыто задним краем переднеспинки. Переднеспинка и щиток с тёмным рисунком. Рисунок переднеспинки часто очень изменчив. На щитке он менее вариабельный, в основном в виде тупоугольного пятна, полной или неполной буквы «W» или пары симметричных треугольных пятен, расположенных вдоль средней линии.
Надкрылья заходят за вершину брюшка. Они отчетливо разделены бороздками на участки — клавус, кориум, кунеус и перепоночку. Кориум с тремя жилками. Кнаружи от кориума располагается эмболиум. Кунеус по внешнему краю отграничен от кориума маленькой вырезкой. У Lygus rugulipennis надкрылья густо опушены, отдельные волоски частично перекрывают друг друга. Перепоночка тёмно-серая, с двумя ячейками: большой и малой.
Ноги довольно длинные и стройные, со сближенными тазиками. Задние бедра немного уже темени. Вершины бёдер с двумя бурыми кольцами, которые могут быть незамкнутыми. Наружная сторона голеней с бурым пятном на колене и еще одним бурым пятном (иногда кольцом) дистальнее первого. Щетинки на голенях чёрные. Вершины голеней и лапки тёмно-бурые. Лапки трех-члениковые.

Рис. 1. Взрослая особь клопика опушённого Lygus rugulipennis [ориг.]:
А) на листе баклажана; Б) на лепестке подсолнечника
Генитальный сегмент самца в форме округленного конуса, лежит на вершине брюшка, не втянут в предшествующие сегменты. Генитальное отверстие на спинной стороне. Эдеагус в покое втянут внутрь. По бокам от него находятся парамеры, которые сильно отличаются друг от друга по форме. Левый парамер крупнее правого. Его гипофиз сплющен в вершинной части, чувствительный бугорок короткий, густо усажен зубчиками (рис. 2, А). Гипофиз правого парамера изогнут почти под прямым углом, тело парамера посредине длины с неглубокой перетяжкой; в основании с сильно торчащим бугорком (рис. 2, Б). Спикула эдеагуса не утолщена к вершине (рис. 2, В).
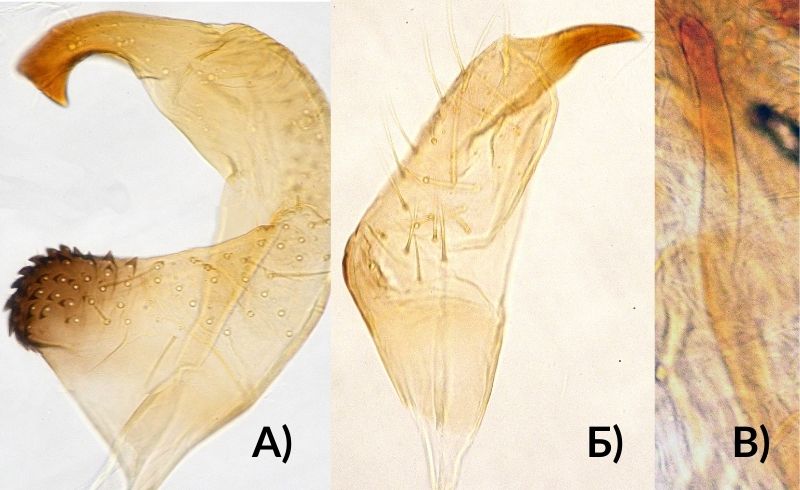
Рис. 2. Генитальный аппарат самца клопика опушённого Lygus rugulipennis [ориг.]:
А) левый парамер; Б) правый парамер; В) спикула эдеагуса
Яйца лигуса беловатые, слегка изогнутые, около 1,7 мм в длину и 0,5 мм в ширину. Самка с помощью зазубренного яйцекладка вставляет каждое яйцо в растительную ткань, чаще при основании листовых черешков. Верхняя часть яйца сплющена в виде крышечки; через неё вылупляется личинка. Эмбриогенез длится в течение 6-14 дней в зависимости от температуры.
Клопы проходят через пять возрастов, прежде чем стать взрослыми. В стадии личинки имеют 3 возраста; они зеленоватые с красными усиками и могут быть ошибочно идентифицированы как тля. По мере взросления, в стадии нимфы (IV и V возраст) у них развиваются четыре подушечки (зачатки) крыльев и пять чёрных пятен на груди и брюшке. Общее время развития всех пяти возрастов колеблется от 15 до 30 дней.
Нимфа IV возраста зеленоватая (рис. 3, А). Нимфа V возраста (рис. 3, Б) с вальковатым телом: длина 3.7-4.4 мм, ширина 1.9-2.2 мм [5]. Голова шириной 1.0 мм, темя – 0.5 мм. Лоб посередине с бурой полоской. Глаза бурые, иногда с рыжим оттенком. Усики 4-х члениковые; 1-й и 2-й членики усиков светлее; 3-й и 4-й – темнее; длина члеников усиков (мм) 0.5, 1.3, 0.8, 0.7. Переднеспинка буро-зеленая, с двумя сближенными черными точками. Длина переднеспинки 0.7 мм, ширина в основании – 1.4 мм. Щиток с бурым рисунком, с двумя расставленными черными точками. Зачатки надкрылий светло-бурые. Брюшко бледно-зеленое. 2-3-й брюшные тергиты вдоль средней линии с округлым желтым пятном перед перитремой пахучей железы. На 3-м тергите расположено одно черное пятно.

Рис. 3. Клопик опушённый Lygus rugulipennis на листе баклажана:
А) нимфа IV возраста; Б) нимфа V возраста (ориг.)
Зимует клопик опушённый Lygus rugulipennis во взрослой стадии преимущественно в лесной подстилке и в ветвях хвойных деревьев, расположенных близко к земле.
Вид унивольтинен в Финляндии [9]: клоп мигрирует на культивируемые поля в середине или конце мая, откладка яиц начинается в конце мая, взрослые особи появляются в конце июля или августе.
В Северной Европе и Северной Америке Lygus rugulipennis является бивольтинным.
В Южной Европе (Италия) клопик развивается в 3-х поколениях [10].
Lygus Rugulipennis доминирует на заболоченных участках, на пойменных лугах малых рек, увлажненных участках, опушках лиственных и сосновых насаждений; наиболее многочислен на залежных участках и невозделываемых полях и участках с рудеральной растительностью; довольно многочислен на посевах сельскохозяйственных культур (люцерна, клевер и проч.) [3].
Имея голарктическое распространение этот высокополифаговой вид широко распространен как на травянистых, так и на древесных дикорастущих растениях и культурах и был зарегистрирован на более чем 320 растениях-хозяевах [11].
На территории европейской части России полевые клопы в качестве кормовых используют 257 видов растений, относящихся к 44 семействам.
В условиях среднерусской лесостепи L. rugulipennis тяготеет к сурепке обыкновенной Barharea vulgaris; в меньшей степени к полыни горькой Artemisia absinthium и злакам [12].
В Швеции L. rugulipennis предпочитает Matricaria perforata, Trifolium medium (L.) и Urtica dioica. в Германии L. rugulipennis отмечается на Senecio vulgaris L., Capsella bursa-pastoris Med., Thlaspi arvense L., Atriplex patula L., Matricaria L. sp., Urtica dioica.
Подчеркивается важная роль сорняков для полевых клопов, которые служат для откладки яиц, а также кормовыми растениями для личинок. Перед зимовкой L.rugulipennis питаются на хвойных или лиственных деревьях, особенно на Corylus avellana L. и Quercus L. sp., а также на Calluna vulgaris (L.) Hill. После зимовки главными кормовыми растениями становятся Vaccinium myrtillus L., Padus avium Mill., Ribes alpinum, Quercus L. и Salix L.
Изменение окраски наблюдается на протяжении жизненного цикла и по сезонам года, наблюдаются также некоторые различия в окраске самцов и самок. Общую закономерность в изменении окраски L. rugulipennis и L. pratensis из Швеции выявил Б. Кулленберг (Kllenberg, 1946). По его наблюдениям молодые имаго светлые, имеют зеленую или желтую окраску. Потом появляется красноватый или красновато-бурый цвет. Перед зимовкой насекомые становятся темными, а весной зимние цвета бледнеют. Самцы светлеют медленнее и иногда остаются ярко окрашенными до самой смерти.
Молодые имаго L. rugulipennis и L. pachycnemis серовато-зеленого цвета. Затем появляются тёмные и буроватые тона, выраженные в большей степени у самцов. Кроме того, у L. rugulipennis в общий цветовой фон включается красноватый цвет (Аглямзянов, 1990). Причины изменчивости черного рисунка до конца не выяснены. Исследованные Э. Вагнером (Wagner, 1949b) темные экземпляры были недавно линявшими особями летнего поколения. Он выделял их как формы, например L. rugulipennis f. obscura. Собранные Р.С. Аглямзяновым (1990) особи L. rugulipennis с «аномально» развитым рисунком на переднеспинке также были летнего поколения.
У L. rugulipennis М. Бонесс (1963) отметил обратную зависимость между площадью чёрного рисунка и температурой. В его исследованиях темный пигмент появлялся у насекомых, которые развивались при сравнительно низких температурах (+15С).
Таблица. Биологические особенности клопика опушённого Lygus rugulipennis при питании стручками зелёной фасоли (температура 25±1°C, относительная влажность воздуха 50-60%, фотопериод 14L:10D) [13]
|
Стадия развития |
Среднее время развития, сут. |
| Яйцо | 10,1 |
| Личинка I | 3,3 |
| Личинка II | 2,6 |
| Нимфа III | 2,8 |
| Нимфа IV | 2,9 |
| Нимфа V | 4,1 |
| Всего (постэмбриональные стадии) | 15,7 |
| Продолжительность жизни самки | 52 |
| Продолжительность жизни самца | 38 |
| Плодовитость | 140-190 |
| Продолжительность откладки яиц | 25-43 |
Клопы Lygus используют действие «рваной раны и 20 мацераций», чтобы вызвать травму: растительные ткани сверлятся или рвутся, а затем слюна вводится в ткань с последующим ферментативным действием (мацерацией) и последующим приёмом клеточного содержимого.
Наиболее важными секретируемыми ферментами, обнаруженными в слюнных железах Lygus rugulipennis, были полигалактуроназа, α-амилаза и щелочные протеиназы [14]. Полигалактуроназа является ферментом, в первую очередь ответственным за пищеварение растительных тканей.
Полигалактуроназа в слюне Lygus представляет собой пектиназу, которая ферментативно переваривает клеточные стенки растений путем солюбилизации (коллоидный процесс проникновения) средней ламели клеточной стенки.
Как только клеточные стенки разрушены, α-амилазы и протеазы помогают в экстра-пероральном переваривании клеточного содержимого.
Трегалазы детоксифицируют аллелохимические вещества растений, подавляя механизмы защиты растений.
Предпочтительное кормление клопов Lygus на меристематических тканях может привести к гормональному дисбалансу, возникающему в результате локализованной травмы в месте питания. Гормональный дисбаланс приводит к изменению вегетативного роста, такому как снижение роста стеблей и увеличение роста боковых ветвей, а также морфологическая деформация плодов или семян.
Из-за важной роли, которую ауксины играют в регуляции абсциссии (сброс органов растения) плодоносящих структур, абсциссия этих структур может происходить в результате разрушения тканей, продуцирующих ауксин-22, в меристематических тканях во время кормления клопом Lygus.
Установлено, что питание Lygus производило меньше абсциссии цветков, когда растения обрабатывали внекорневым ауксином и нафталин уксусной кислотой [15].
Когда лигусные клопы присутствуют в большом количестве, налив семян и созревание могут быть уменьшены почти на 100%. Нимфы и взрослые особи атакуют стебли и репродуктивные части растений (рис. 3). Во время кормления клопы-лигусы могут вводить токсичную слюну в растение и вызывать отмирания и опадение семенных структур с растения. Даже умеренное кормление может привести к преждевременному сбрасыванию почек, деформированным семенам и снижению жизнеспособности семян.

Рис. 4. Повреждения от клопа, баклажаны осенние (второй оборот) Бьенсе. Фото от 2.08.21 elena.tchecheneva.
 Рис. 5. Завязь (горох) баклажана, готовая отвалиться [ориг.] Повреждение клопом нанесено в стадии бутонизации.
Рис. 5. Завязь (горох) баклажана, готовая отвалиться [ориг.] Повреждение клопом нанесено в стадии бутонизации.
Пестицидные обработки должны быть отложены до тех пор, пока не будет завершено вылупление из яиц, но до того, как нимфы достигнут IV-V возраста.
В Аризоне были протестированы 11 пестицидов [16], но ни один из них не был активен против взрослых Lygus, тогда как некоторые химические вещества были очень эффективны на всех возрастах нимф. Orthene® и Vydate® показали хорошие результаты против Lygus, но были недостаточно эффективны.
Наиболее эффективным инсектицидом против Lygus был флоникамид (Teppeki), разрабатываемый FMC. Этот инсектицид имел лучший контроль над нимфами Lygus, был самым высокопроизводительным и требовал на одну обработку меньше, чем другие наиболее эффективные соединения. Среди новых химических веществ для контроля Lygus был фипронил (Regent®), который работал немного лучше, чем Vydate, но был не так эффективен, как Orthene. Из неоникотиноидов динотефуран (Valent) показал хорошую активность. Производительность спиромезифина (Оберон®) была похожа на динотефуран, но нуждалась в одном распылении меньше.
В полевых исследованиях на землянике против местного вида Lygus hesperus (Калифорния) было показано влияние флоникамида (Beleaf 50 SG) по норме 199.6 г препарата/га. Установлено, что препарат умеренное подавление численности как имаго, так и нимф клопа при двукратной обработке в течение 2-х недель [17]. В этих же исследованиях препараты на основе тиаметоксама и фенпропатрина проявили низкую эффективность.
Известно, что в Европе на Lygus rugulipennis паразитируют три вида Peristenus (Hymenoptera: Braconidae: Euphorinae) — Peristenus rubricollis (Thomson, 1891), Peristenus digoneutis Loan, 1973 и Peristenus stygicus Loan, 1973 [18].
Виды Peristenus откладывают яйца в молодых личинок мирид. Взрослые личинки паразитоида появляются либо в V стадии клопа, либо во взрослых особях, где и окукливаются. Развитие паразитоидов довольно длительное. Например, у P. rubricollis требуется 47 дней от откладки яйца до окукливания. Наиболее высокий уровень паразитирования наблюдается во второй половине июля и первой половине августа. Жизненный цикл паразитоида, по-видимому, синхронизирован с развитием Lygus.
В Канаде Lygus rugulipennis, имеет два поколения нимф в год, первое из которых обычно характеризуется более низким уровнем паразитации (4-27%). В среднем уровень паразитизации у второго поколения нимф выше 32%, изредка достигая более 60%. Peristenus digoneutis доминирует в местах произрастания клевера, тогда как на ромашковых насаждениях P. stygicus и P. digoneutis появляются с одинаковой частотой [19].
Также было зарегистрировано, что на яйцах Lygus rugulipennis паразитируют Anaphes fuscipennis Haliday, 1833 (Hymenoptera: Mymaridae) [20].
По вопросам биологической защиты растений в теплицах, разработки проектов защиты, внедрения IPM метода обращайтесь к технологам компании Биозащита.
[1] Die Miridae Hahn, 1831, des Mittelmeerraumes und der Makaronesischen Inseln (Hemiptera, Heteroptera). E Wagner and HH Weber, 1970.
[2] Aglyamzyanov R. Revision der paläarktischen Arten der Gattung Lygus Hahn, 1833 (Heteroptera: Miridae) // Dissertation. http://doi.org/10.25358/openscience-4163 // Entomologische Zeitschrift mit Insekten-Börse. – 2009. – Vol.119. – No. 6. – P. 249-276.
[3] Демченко Н. Ю. Фауна и экология полужесткокрылых насекомых рода Lygus Hahn (Heteroptera, Miridae) европейской части России / Автореферат диссертации на соискание ученой степени кандидата биологических наук. – Воронеж. – 2004. – 20 с.
[4] Пучков В. Г. Главнейшие клопы-слепняки – вредители сельскохозяйственных культур / Киев: Наукова думка. – 1966. – 172 с.
[5] Poppius B. Eine neue Lygus-Art aus Finland // Meddelanden af Societas pro Fauna et Flora Fennica. via Catalogue of Life. – 1911. – T. 37. – S. 96-98.
[6] Schwartz M. D., Foottit R. G. Revision of the Nearctic Species of the Genus Lygus Hahn, with a Review of the Palaearctic Species (Heteroptera: Miridae) // Memoirs on Entomology, International. Gainesville, Florida, U. S. A.: Associated Publishers. – 1998. – Vol. 10. – 428 p.
[7] Kerzhner I. M., Josifov M. Family Miridae Hahn, 1833. Cimicomorpha II. / In: B. Aukema, Ch. Rieger (eds.). Catalogue of the Heteroptera of the Palaearctic Region // Amsterdam, The Netherlands: The Netherlands Entomology Society/ – 1999. – Vol. 3. – 577 p.
[8] Винокуров Н. Н., Канюкова Е. В., Голуб В. Б. Каталог полужесткокрылых насекомых (Heteroptera) Азиатской части России / Новосибирск: Наука. – 2010. – 320 с.
[9] Varis A.L. The biology of Lygus rugulipennis Popp. (Het. Miridae) and the damage caused by this species to sugar beet // Annales Agriculturae Fenniae. – 1972. – Vol. 11. – N. 156. – S. 1-56. http://urn.fi/URN:NBN:fi-fe2014102745943
[10] Pansa M.G., Guidone L., Tavella L. Distribution and abundance of nymphal parasitoids of Lygus rugulipennis and Adelphocoris lineolatus in northwestern Italy // Bulletin of Insectology 2012. – Vol. 65. – N. 1. – P. 81-87,
[11] Holopainen J.K. Host plant preference of the tarnished plant bug Lygus rugulipennis Popp. (Heteroptera, Miridae) // J. Appl. Entomol. – 1989. – Vol. 107. – P. 78-82.
[12] Демченко Н.Ю., Голуб В.Б. Экологические особенности Lygus pratensis L. и L. rugulipennis Popp. (Heteroptera, Miridae) в условиях лесостепи и севера степной зоны европейской части России // Состояние и проблемы экосистем среднерусской лесостепи. — Воронеж, 2003. (Тр. учеб.-науч. центра Воронеж, гос. ун-та «Веневитиново»; вып. 16). — С. 44-51.
[13] Salerno G., Frati F., Conti E., Bin F. Influence of different diets and oviposition substrates on Lygus rugulipennis biology (Heteroptera: Miridae) // Eur. J. Entomol. – 2007. – Vol. 104. – P. 417-423. – DOI: 10.14411/eje.2007.061
[14] Laurema S., Varis A.-L., Harri M. Studies on enzymes in the salivary glands of Lygus rugulipennis (Hemiptera, miridae) // Insect Biochemistry. – 1985. – Vol. 15. – Is. 2. – P. 211-224. – doi.org/10.1016/0020-1790(85)90010-1
[15] Fisher E.H., Riker A.J., Allen T.C. Bud, blossom, and pod drop of canning string beans reduced by plant hormones // Phytopathology. – 1946. – Vol. 36. – P. 504-523.
[16] Barkley V., Ellsworth P.C. Search for Effective Chemical Controls for Lygus Bugs and Whiteflies in Arizona Cotton // Arizona Cotton Report (P-138). – 2004. – P. 186-205. – www.researchgate.net/publication/253652617
[18] Joseph S.V., Bolda M. Efficacy of insecticides against Lygus hesperus Knight (Hemiptera: Miridae) in the California’s Central Coast strawberry // International Journal of Fruit Science. – 2016. – Vol. 16. – Is. supl: Proceedings of the 2015 North American Strawberry Symposium Submit an article Journal homepage. – P. 178-187. – doi.org/10.1080/15538362.2016.1219293
[19] Haye T. Studies on the ecology of European Peristenus spp. (Hymenoptera: Braconidae) and their potential for the biological control of Lygus spp. (Hemiptera: Miridae) in Canada // Aus dem Zoologischen Institut –Zoophysiologie. – Dissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultät der Christian-Albrechts-Universität zu Kiel vorgelegt von. – Kiel, Mai 2004
[20] Bilewicz-Pawi´nska T. Plant bugs (Heteroptera, Miridae) and their parasitoids (Hymenoptera, Braconidae) on cereal crops // Polish Ecol. Studies. – 1982. – Vol. 8. – P. 113-191.
Статью для блога Биозащиты подготовил кандидат биологических наук, старший научный сотрудник, акаролог Мешков Юрий Иванович.




Оставить комментарий